
Tuberculosis as an infectious pathology of immune system
Keywords:
tuberculosis, immunity, phagocytosis, vaccinesAbstract
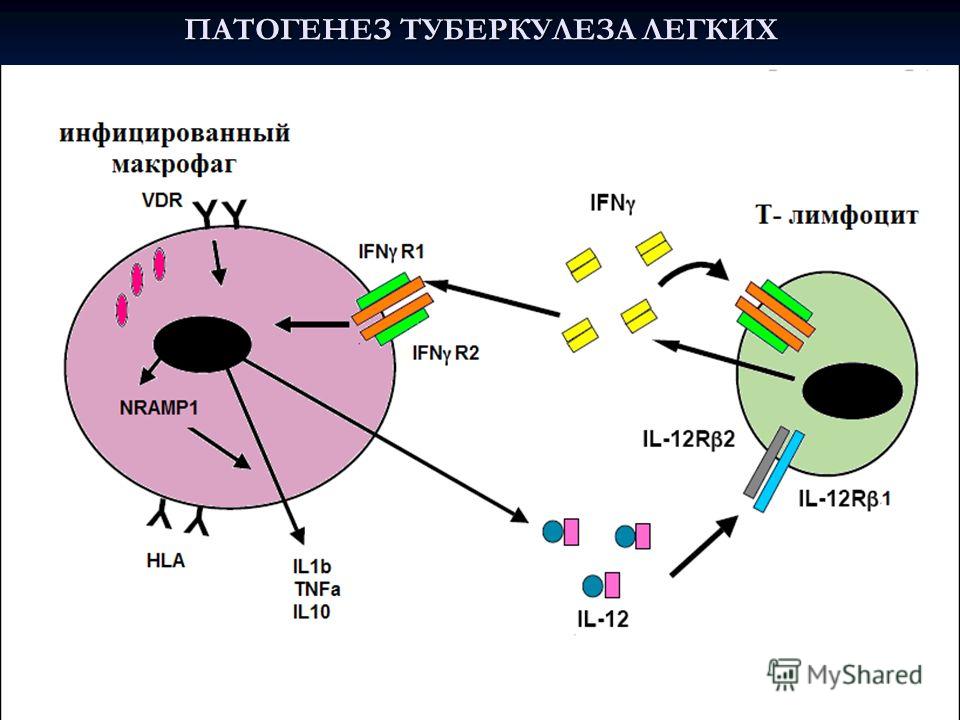
As a result of years’ research of the many research groups around the world able to understand the reason why it will be impossible to create really effective vaccine for the prevention of tuberculosis infection in the near future. The main reason for the impossibility creating such vaccine is an intracellular nature of tuberculosis. In fact, TB is a pathology of the immune system. Mycobacterium tuberculosis persist within macrophages and thereby inhibit the process of phagocytosis completion and digesting the contents of phagosome. The destruction of the lysosomal membrane inside macrophages is blocked by changing the pH in lysosomes. For the presence of lytic activity for most lysosomal enzymes require need acidic environment. Mycobacteria are also getting into the lysosomes of macrophages start to rapidly hydrolysis for urea by urease to form ammonia. Wherein pH in the medium changes to alkaline, this inactivates enzymes and stabilizes lysosomal membrane. Thus mycobacterium prevent lysosome collapse at inactivated lysosomal enzymes and do not allow them to complete macrophage digestion phase by transition lysosomal to phagosomal stage. Stop phagocytolysis process leads to imbalance of the host immune system. Increasing the number of infected macrophages sensitized to Mycobacterium tuberculosis antigens, leading to constant hyperfunction of cellular immunity, particularly enhanced immune response to cell wall components of mycobacteria, induction high titers of interferon-gamma in response to a stimulus, a sharp jump IL-2 titers and TNF-α , IFN-γ specific activation CD8 + CTL. Need also focus attention on the main differences from the MBT and human BCG, that is well growth in the human body, persists along host life, but does not cause active TB (except in patients with HIV/AIDS). After MBT cell destruction in the environment gets some additional high allergenic antigens, such as 85B, ESAT6, Rv2660c, HyVaC 4 (Ag85B and TB10.4.). These antigens to provide high adhesion and allergenicity of human strains M. tuberculosis. Most allergens that cause obvious signs of active tuberculosis are the antigens ESAT6 and CFP10. Such protein antigens can be called endotoxins. Also to pathogenicity factors include cord-factor, it main component is a polysaccharide-mycolic complex from cell wall (Figure 2) containing ftiolic and mycolic acid - to ensure the stability of mycobacteria to lysosomal enzymes. Currently available diagnostic tools tuberculin preferably contain the above components of the cell wall and differences (from BCG) allergens ESAT6 and CFP10 []. Currently well established that the virulence of M. tuberculosis, mainly responsible genes encoding antigens ESAT-6 and CFP10. When comparing the genomic sequence of M. tuberculosis with attenuated M. bovis BCG was detected genomic deletion of the three sites in the vaccine strain (RD1, RD2, RD3). BCG vaccine strain genome stripped areas in the RD1, encoding mycobacterial antigens ESAT-6 and CFP-10 present in virulent strains of M. tuberculosis. Many researchers believe that mutations in genomic regions RD1, encoding mycobacterial antigens ESAT-6 and CFP-10, occurred in the process of creating a BCG strain. It remains not examine the question of whether all strains of M. bovis other than BCG have antigens ESAT-6 and CFP-10, and whether they depend on the degree of virulence of the mycobacteria strains. About a third of the population is infected with the MBT. Tuberculosis statistics show that out of every 100 man infected MTB, only 10 appear open clinical forms. In the remaining patients, positive skin test and/or gamma-interferon test, clinical symptoms of tuberculosis never does occur, and no signs of sensitization other than to MBT antigens and presence ESAT6 - antibodies in the blood. Thus, if the focus is not on the infection, but on the prevention of tuberculosis reactivation, can significantly reduce the number of cases with clinical manifestations. There have been recent publication comparing the immunity of patients with open clinical forms tuberculosis and without clinical symptoms, but ESAT6 - test-positive. One of the rational ways for helps to MTB - infected macrophages is the simultaneous use of urease inhibitors and simultaneously use selective activators of antibacterial complete phagocytosis. For the latter group, some authors include also histone deacetylase inhibitors (HDAi). The use of such inhibitors in the latter case will mass increase number reading frames in the macrophages genome and leads to stormy expression phagocytosis activators, that blocked by MBT. These inhibitors include valproic acid and trichostatin. Research in this area only started, and the expectations are very high. Another activator phagocytosis with very similar action mechanisms is the vitamin D3 - ergocalciferol. In a variety experiments shows that the soluble derivatives of vitamin D3 inoculation to the culture of MBT - infected macrophages leads to the completion phagocytosis and complete digestion of the MBT. The disadvantage of this method is the need to maintain a concentration of vitamin D3, which is quite toxic to the human body as a whole. Accordingly, a new form vitamin D3 is to be administered directly to the places where many infected macrophages, i.e. as an aerosol through the lungs. Also pay attention to the fact that, earlier for purpose combating tuberculosis the urease inhibitors have not been used, although quite a lot of well-known non-toxic compounds anti -urease activity. Thus, the most promising way to prevent tuberculosis reactivation in humans with positive test specimens and humans in remission following chemotherapy is to provide an aerosol preparation containing both urease inhibitor, activator phagocytosis vitamin D3 and histone deacetylase inhibitor. The use of such aerosol once a week will greatly reduce the number of macrophages with incomplete phagocytosis and prevent the background to tuberculosis with clinical open forms. This disease, like tuberculosis, prevention is better than cure, especially with the emergence of M. tuberculosis multiresistant strains.
References
Hesseling A.C. Resistant Mycobacterium bovis Bacillus Calmette-Guerin disease: implications for management of Bacillus Calmette-Guerin Disease in human immunodeficiency virus-infected children/ A.C. Hesseling, H.S. Schaaf, T. Victor et al. // Pediatr Infect Dis.- 2004.- Vol.23.-P.476–479
Hatherill M. Prospects for elimination of childhood tuberculosis: the role of new vaccines/ M. Hatherill //Arch Dis Child.-2011.- Vol.96.- P.851–856.
Rodrigues L.C. Effect of BCG revaccination on incidence of tuberculosis in school-aged children in Brazil: the BCG-REVAC cluster-randomised trial/ L.C. Rodrigues, S.M. Pereira, S.S. Cunha, et al. // Lancet.- 2005.-Vol. 366.- P. 1290-1295.
Chen L. Single intranasal mucosal Mycobacterium bovis BCG vaccination confers improved protection compared to subcutaneous vaccination against pulmonary tuberculosis/ L. Chen, J. Wang, A. Zganiacz, Z. Xing. // Infect.Immun.- 2004.-Vol. 72.- P.238–246.
Barclay W.R. Protection of monkeys against airborne tuberculosis by aerosol vaccination with bacillus Calmette-Guerin/ W.R. Barclay, W.M. Busey, D.W. Dalgard et al. // Am Rev Respir Dis.- 1973.- Vol. 107.- P.351–358.
Flynn J. L. Immunology of tuberculosis/ J. L. Flynn, J. Chan. //Annual Review of Immunology. — 2001. — Vol. 19. — Р. 93—129
Maianskii A. N. Tuberculosis/ A.N. Maianskii // Immunology.- 2001.-N. 2.- P. 53—63
Reinout van Crevel. Innate Immunity to Mycobacterium tuberculosis/ R. Crevel, T. H.M. Ottenhoff, Jos W.M. van der Meer. // Clinical Microbiology Reviews.- 2002.- Vol. 15, N. 2.-P. 294—309.
Chernushenko E.F. Cytokines in the evaluation of the immune system in patients with pulmonary tuberculosis / E.F. Chernushenko, L.P. Kadan, O.R. Panasyukova et al. // Ukrainian pulmonology journal. - 2010. - № 2. - p. 39-43
http://images.myshared.ru/9/935683/slide_36.jpg
Reinout, van Crevel. Innate Immunity to Mycobacterium tuberculosis / van Crevel Reinout, Tom H. M. Ottenhoff, Jos W. M. van der Meer // Clinical Microbiology Reviews. - 2002. - V. 15. - №. 2. - P. 294—309.
Reinout, van Crevel. Innate Immunity to Mycobacterium tuberculosis / van Crevel Reinout, Tom H. M. Ottenhoff, Jos W. M. van der Meer // Clinical Microbiology Reviews. - 2002. - V. 15. - №. 2. - P. 294—309.
Chan, J. Killing of virulent Mycobacterium tuberculosis by reactive nitrogen intermediates produced by activated murine macrophages / J. Chan, Y. Xing, R. S. Majliozzo, B. R. Bloom // J. Exp. Med. - 1992. - № 175. - P. 1111–22.
Leemans, J. C. Macrophages play a dual role during pulmonary tuberculosis in mice / J. C. Leemans, T. Thepen, S. Weijer, S. Florquin, T. V. D. Poll // J. Infect. Dis. - 2005. - № 191. - P. 65–74.
Grode, L. Increased vaccine efficacy against tuberculosis of recombinant Mycobacterium bovis bacille Calmette-Guerin mutants that secrete listeriolysin / L. Grode, P. Seiler, S. Baumann et al. // J Clin Invest. - 2005. - № 115. - P.2472–9.
Meena, L. S. Rajni. Survival mechanisms of pathogenic Mycobacterium tuberculosis H37Rv / L. S. Rajni Meena // FEBS J. - 2010. - № 277. - P. 2416-27.; PMID:20553485; http://dx.doi.org/10.1111/j.1742-4658.2010.07666.x
Serbina, N. V. CD8_ CTL from lungs of Mycobacterium tuberculosis-infected mice express perforin in vivo and lyse infected macrophages / N. V. Serbina, C. C. Liu, C. A. Scanga, and J. L. Flynn // J. Immunol. - 2000. - №165. - P. 353–63.
Wards, B. J. An esat6 knockout mutant of Mycobacterium bovis produced by homologous recombination will contribute to the development of a live tuberculosis vaccine / B. J. Wards, G. W. de Lisle, D. M. Collins // Tuber Lung Dis. - 2000. - №80. - P. 185-9. PMID:11052907; http://dx.doiorg/10.1054/tuld.2000.0244
Ravn, P. Human T cell responses to the ESAT-6 antigen from Mycobacterium tuberculosism/ P. Ravn, A. Demissie, T. Eguale, H. Wondwosson, D. Lein, H. A. Amoudy et al. // J Infect Dis. - 1999. - № 179. - P. 637-45.; PMID:9952370; http:// dx.doi.org/10.1086/314640
Pym, A.S. Recombinant BCG exporting ESAT- 6 confers enhanced protection against tuberculosis / A. S. Pym, P. Brodin, L. Majlessi, R. Brosch, C. Demangel, A. Williams et al. // Nat Med. - 2003. - V. 9. - P. 533-539.; PMID:12692540; http://dx.doi.org/10.1038/nm859
Pallen, M. J. The ESAT-6/WXG100 superfamily – and a new Gram-positive secretion system? / M. J. Pallen // Trends Microbiol. - 2002. - V. 10. - P. 209-12.; PMID:11973144; http://dx.doi.org/10.1016/S0966-842X(02)02345-4
Arend, S. A. Double-blind randomized Phase I study comparing rdESAT‑6 to tuberculin as skin test reagent in the diagnosis of tuberculosis infection / S. A. Arend, W. P. Franken, H. Aggerbeck et al. // Tuberculosis.— 2008. — V. 88. — P. 249–61..
Magdalena, J. Specific differentiation between Mycobacterium bovis BCG and virulent strains of the Mycobacterium tuberculosis Complex/ J. Magdalena, Ph. Supply, C. Locht // Clin. Microbiol. - 1998. - V. 36. - № 9. - P. 2471–76.
Harboe, M. Evidence for occurrence of the ESAT-6 protein in Mycobacterium tuberculosis and virulent Mycobacterium bovis and for its absence in Mycobacterium bovis BCG / M. Harboe, Т. Oettinger, H. G. Wiker, I. Rosenkrands, P. Andersen // Infect. Immun. - 1996. - V. 64. - Р. 16–22.
Schaible, U. E. Cytokine activation leads to acidification and increases maturation of Mycobacterium avium-containing phagosomes in murine macrophages / U. E. Schaible et al. // J. Immunol. - 1998. - V. 160. - № 3. - P. 1290–96.
Reyrat, J. M. The urease locus of Mycobacterium tuberculosis and its utilization for the demonstration of allelic exchange in Mycobacterium bovis bacillus Calmette-Guérin / J. M. Reyrat, F. X. Berthet, B. Gicquel // Proc. Natl. Acad. Sci. U. S. A. - 1995. - Т. 92. - № 19. - P. 8768–72.
Gordon, A. H. Ammonia inhibits phagosome-lysosome fusion in macrophages / A. H. Gordon, P. D’Arcy Hart , M. R. Young // Nature. - 1980. - Т. 286. - № 5768. - P. 79–80.
Grode, L. Safety and immunogenicity of the recombinant BCG vaccine VPM1002 in a phase 1 open-label randomized clinical trial / L. Grode, C. A. Ganoza, C. Brohm, J 3rd Weiner, B. Eisele, S. H. Kaufmann // Vaccine. - 2013. - V. 31. - P. 1340-48.
Desel, C. Recombinant BCG ΔureC hly+ induces superior protection over parental BCG by stimulating a balanced combination of type 1 and type 17 cytokine responses / C. Desel, A. Dorhoi, S. Bandermann, L. Grode, B. Eisele, S. H. Kaufmann // J Infect Dis . - 2011. – Vol. 204 – P. 1573-84.
Farinacci, M. The recombinant tuberculosis vaccine rBCG ΔureC::hly(+) induces apoptotic vesicles for improved priming of CD4(+) and CD8(+) T cells/ M. Farinacci, S. Weber, S. H. Kaufmann // Vaccine. - 2012. - V. 30. - P. 7608-14.
Study to Evaluate Safety and Immunogenicity of VPM1002 in Comparison With BCG in Newborn Infants in South Africa. [http:// clinicaltrials.gov/ct2/show/NCT01479972?term=vpm1002&rank=2].
Kaufmann, S. H. Recombinant live vaccine candidates against tuberculosis / S. H. Kaufmann, M. Gengenbacher // Curr Opin Biotechnol. - 2012. - V. 23. - P. 900-907.
Hoft, D. F. A new recombinant bacille Calmette-Guérin vaccine safely induces significantly enhanced tuberculosis-specific immunity in human volunteers / D. F. Hoft, A. Blazevic, G. Abate, W. A. Hanekom, G. Kaplan, J. H. Soler, F. Weichold, L. Geiter, J. C. Sadoff, M. A. Horwitz // J Infect Dis. - 2008. - № 198. - P. 1491-501.
Arbues, A. Construction, characterization and preclinical evaluation of MTBVAC, the first live-attenuated M. tuberculosis-based vaccine to enter clinical trials / A. Arbues, J. I. Aguilo, J. J. Gonzalo-Asensioet al. // Vaccine. - 2013. - V. 31. - P. 4867-73.
Nambiar, J. K. Protective immunity afforded by attenuated, PhoP-deficient Mycobacterium tuberculosis is associated with sustained generation of CD4+ T-cell memory / J. K. Nambiar, R. Pinto, J. I. Aguilo, et al.// Eur J Immunol. - 2012. - № 42. - P. 385-92.
Martin, C. The live Mycobacterium tuberculosis phoP mutant strain is more attenuated than BCG and confers protective immunity against tuberculosis in mice and guinea pigs / C. Martin, A. Williams, R. Hernandez-Pando, et al. // Vaccine. - 2006. - V. 24. - P. 3408-3419.
WHO Tuberculosis Reviewed March 2016. [http://www.who.int/mediacentre/factsheets/fs104/en/]
Kasprowicz, V. O. Diagnosing Latent Tuberculosis in High-Risk Individuals: Rising to the Challenge in High-Burden Areas / V. O. Kasprowicz, G. Churchyard , S. D. Lawn , S. B. Squire, A. Lalvani // J Infect Dis. - 2011. - V. 4. - № 204. - P. 1168–78.
Khonina, N.A. Features of immunity in patients with various forms of pulmonary tuberculosis / N.A. Khonina, SD Nikonov SV Shpilevsky et al. // Problems of tuberculosis. - 2000. - № 1. – P. 30-32.
Tsao, T. C. Y. Increased TNF-α, IL-1β and IL-6 levels in the bronchoalveolar lavage fluid with the upregulation of their mRNA in macrophages lavaged from patients with active pulmonary tuberculosis / T. C. Y. Tsao et al. //Tubercle and lung disease. – 1999. – Vol. 79. – №. 5. – С. 279-285.
Olobo, J. O. Circulating TNF‐α, TGF‐β, and IL‐10 in Tuberculosis Patients and Healthy Contacts / J. O. Olobo and et al. //Scandinavian journal of immunology. – 2001. – V. 53. – №. 1. – С. 85-91.
Magner, W. J. Activation of MHC class I, II, and CD40 gene expression by histone deacetylase inhibitors/ W. J. Magner and et al. //The Journal of Immunology. – 2000. – V. 165. – №. 12. – С. 7017-7024.
Xie, L. Proteome-wide lysine acetylation profiling of the human pathogen Mycobacterium tuberculosis / L. Xie, X. Wang, J. Zeng, et al. // The international journal of biochemistry & cell biology. - 2015. - № 59. - P. 193-202.
Campbell, G. R. Vitamin D inhibits human immunodeficiency virus type 1 and Mycobacterium tuberculosis infection in macrophages through the induction of autophagy / G. R. Campbell, S. A. Spector // PLoS Pathog. – 2012. – V. 8. – №. 5. – P. e1002689.
Chandra, G. Effect of vitamin D3 on phagocytic potential of macrophages with live Mycobacterium tuberculosis and lymphoproliferative response in pulmonary tuberculosis /G. Chandra, P. Selvaraj, M. S. Jawahar, V. V. Banurekha, P. R. Narayanan //Journal of clinical immunology. – 2004. – V. 24. – №. 3. – P. 249-257.
Phillips, K. Antibacterial action of the urease inhibitor acetohydroxamic acid on Helicobacter pylori / K. Phillips, D. J. Munster, R. A. Allardyce, P. F. Bagshaw //Journal of clinical pathology. – 1993. – V. 46. – №. 4. – P. 372-373.
Downloads
How to Cite
Issue
Section
License
Copyright (c) 2020 Annals of Mechnikov's Institute

This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 3.0 Unported License.


{kind=link}